第 7 回の講義(1 月 31 日)の質問への回答です。
すみません。18:00 ちょっと加筆しました。
細胞分化,核移植実験
(加藤君) ガードンの実験でできたオタマジャクシはクローンですか。
(三好君) 核移植実験によってつくられたオタマジャクシと普通に交配されてできたオタマ
ジャクシは,何ら違いはないんですか?
(答) 加藤君の言うとおり,ガードンが核移植実験で作ったのは体細胞クローンです。
つまり,みんなもよく知っているヒツジのドリーと同じです。なので,まったく同じと
言うわけではないでしょうね。ただ,クローンカエルの方は,それ以後,遺伝子の状態
とか,それほど詳しくは調べられていないと思うので,どこがどう違うか…は詳しくは
わからないと思います。
発生と遺伝
(長沼君) 遺伝子の変異で頭から触角が生えたハエの写真を見ました。同様のことが人間でも
起こりますか(例えば人間の胸に腕を生やすなど)。また,異常なところに生えてきた体は
機能しますか。
(答) それは重要な質問です。頭から脚の生えたヒト…なんて見たことありませんよね。
そういう異常が自然には起こりにくい理由が一つあります。脊椎動物では,ある目的を達成
するための反応経路が 2 つ以上重複している場合が多いのです。それぞれの経路を進める因子
(転写調節因子やシグナル分子,酵素など)は別々なので,それらの因子のどれか一つの機能
が損なわれて,一つの経路が止まったとしても,もう一つの経路が働いていれば,目に見えた
異常はあらわれないのです。そういうわけで,ヒトの頭に脚が生えたなんていうことはない
のです。遺伝子の突然変異は,ハエでもヒトでも,ある程度の頻度で起こります。そこの違い
ではありません。
脊椎動物の進化の過程では,全ゲノム規模の遺伝子重複が 2 回(魚類ではさらにもう 1 回)
起こって,同じ(あるいはよく似た)配列の遺伝子が 3 倍とか 4 倍 (魚類では 7 倍とか 8 倍)
に増えているのです。もともとゲノム中に 1 個だった遺伝子が 2 個になると,片方は壊れても
いいし,別の機能をもつように変化してもいい・・・自由になります。それが新しい遺伝子を
生み出し体の構造を複雑化させる原因になったとも考えられています・・・。でも,2 個になって
相当時間がたっても,遺伝子はそう簡単に変化するものではありませんから,少なくとも部分的には
同じような機能をもっていて,同じ細胞で発現することが多いのです。こういう状態になっている
ので,ショウジョウバエと同じような実験で 1 個の遺伝子に突然変異を起こしても,何の影響も
なく元気に一生を過ごすという結果になる場合が非常に多いのです。言ってみれば,遺伝子重複
によって安全装置ができたようなものですね。ただ,ヒトでも,手足の指が 6 本あったり…と
いう程度のことは起こります。
(井上君) Antennapedia 変異体のハエは触角がありませんが,生きていくことに不自由はない
のでしょうか。
〜似たような趣旨の質問: 三輪さん,小梶君
(答) 触角は大事な感覚器官ですから,ないと,とても生活に困ると思います。自然界では
生き延びられないかも知れませんね。
(深澤君) アンテナペディア変異体では,触角部分に脚ができましたが,他に同じような変異が
生じるものはありますか。
(答) 「同じような」がどういう意味なのか,深澤君の真意を誤解しているかも知れませんが
Antennapedia と同じように「体のある部分の構造が,別の部分の構造に置き換わるような
突然変異」のことを「ホメオティック変異 homeotic mutation」といいます。インパクトの
ある写真でよく知られている例では,平均棍が翅になる Ultrabithorax があります(下図)。
昆虫の胸部は 3 つの体節でできています。前胸部には前脚があります。中胸部は大きく発達
していて前翅と中脚があります。後胸部にはトンボやチョウなどでは後翅と後ろ脚があります。
しかしハエやアブ,カなどの双翅目昆虫では後翅が平均棍という構造に変化しています。説明
が長くなってしまいましたが,Ultrabithorax がまったく機能しない変異体では,後胸部が
そっくりそのまま中胸部に変化してしまうので,本来なら平均棍が生えるべきところに前翅
が生え,本来後ろ脚が生えるべきところに中脚が生えてしまいます。

ウニの胚発生
(Yang 君) Hans Driesch の実験結果について,4 細胞期のとき,集まっている 4 つの細胞は
それぞれ互いに感じられるので,予定運命のとおりに発生するのだろう。これもシグナル分子の
やり取りに関係があるか?
(答) いい質問です。バラバラにすればそれぞれが 1 匹の完全な幼生になれる能力をもつのに,
4 個の細胞がくっついていれば,個々の細胞は体の 4 分の 1 を作るだけ。このとき,細胞は
隣に細胞がくっついていることを何らかの方法で感知しているはずです。それは,たぶん
ホルモンや細胞増殖因子のようなシグナル分子ではないと思います。どうやってそれを感知
しているかは謎です。現時点では明快な答えはないと思います。
(三輪さん) ウニの細胞を半分にするとき,大きさが異なっても正常に発生できるのか。
(谷島君) 下図のように,動物極側と植物極側の量を不均一に切断すると,動物極側はプルテウス
幼生になりますか?
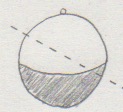
(答) 大きさが少々異なっていても,発生は正常に進みます。また,谷島君の図と同じくらいの
斜めのカットだと,動物極側がプルテウス幼生になることはないだろうと思います。
(小田君) ウニの 4 細胞期にバラバラにしたものが,もう一度分裂したあとまたバラバラに
したらどうなりますか。
(答) それはいい質問ですね。下図の左上が 4 細胞期。そのまま細胞分裂が進んで 8 細胞期に
なったのが右上。 4 細胞胚の割球を一つ単離したのが左下です。もう一度分裂したときに
本来の方向にピンクの細胞と水色の細胞に分裂すれば,それは 8 細胞胚の 8 個の細胞をバラバラ
にしたときと同じ結果になることが期待されますよね。ただ,問題は,単離した 4 細胞胚の割球
が,本来の方向に分裂するのか(A),それとも分裂の向きが変わるようなことがあるのか(B)
どっちなんだろうかってことですよね。これは私にはわかりません。そこで,ウニの専門家である
愛媛大学の高田裕美先生にお尋ねしたところ,下記のようなお返事をいただきましたので転載
します。
【高田先生のお返事】
通常、丁寧に解離を行った場合、1/4 割球の次の分裂方向は正常な場合と同じになります。
動植半球を分ける方向になりますから、そのまま培養すれば、動物半球由来の割球からは
胚壁が薄い中空の永久胞胚が、植物半球由来の割球からは胚壁の厚い細胞が詰まった原腸胚の
出来損ないのような胚が生じます。植物半球由来の 1/8 胚は、形態がいろいろで植物極側が
外側にはみ出している場合もあります。植物半球由来の部分胚は、色素細胞が分化して赤い色が
でてくるので、動物割球由来の部分胚と容易に見分けがつきます。単離した割球を混ぜて培養
すると、互いに癒合して巨大な細胞の塊になったり、何事もなかったかのような普通の胚に
なったりすることもあります。
上手に単離できた 1/4 細胞から生じた 1/8 植物割球からは小割球ができますし、その小割球
は、うまくすると次の卵割で大小割球と小小割球にわかれます。
ただし、バッチによっては、小割球そのものができない場合もあります。これは割球を単離
しなくても受精膜をとっただけでも起こります。シーズンの終わりかけによく見られる現象で、
よく泣かされました。
ちなみに動物半球の細胞は、もともとそれほどきちんと縦横に分裂するわけではありません。
まったく何もしていない正常胚でも斜めに分裂する場合もあります。
厳密なことを言えば、おそらく4細胞期ですでに各割球は等価ではありません。Horstadius の
原図でも微妙に 4 個体のプルテウス幼生の形態が異なっています。 2008 年にハスノハカシパン
で小南先生が論文を出していますが、どうも受精時に背腹(口側反口側)がある程度決まる
ようです。ただし、調節性が高いため、あまり顕著な差となって現れないようです。
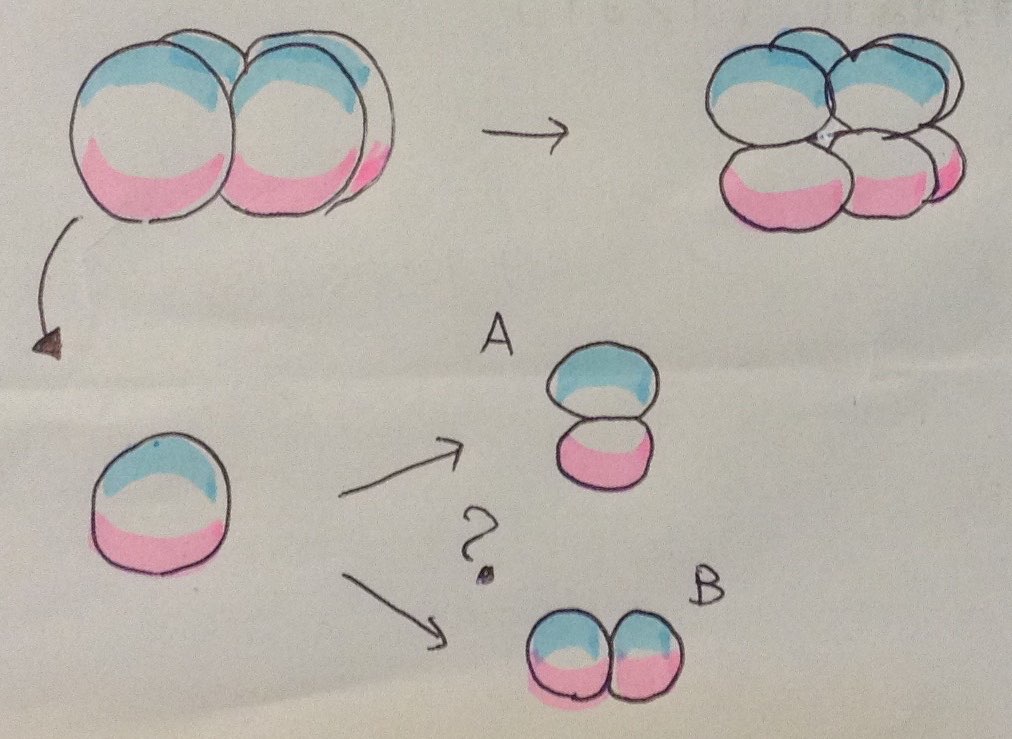
(岡崎君) ウニ胚の割球解離実験でできた 4 分の 1 の大きさのプルテウス幼生をそのまま育てると
成体になっても 4 分の 1 の大きさなのですか。
(答) 4 個の幼生は,最初のうちは小さいですが,エサを食べ始めたらやがては大きく成長して
正常に発生した幼生と区別がつかなくなります。一卵性の双子の一人一人の体の大きさ,私たち
とちっとも違わないでしょう?
(有吉さん) 16 細胞期に,大・中・小で分けると Dsh タンパク質が少量存在する大割球は
どうなりますか?
(答) いい質問です。正常発生では,小割球から骨片形成細胞が分化します。一方,大割球
からは原腸などが生じます。小割球由来の細胞は原腸に分化しないし,大割球からは骨片形成
細胞は分化しません。このような胚から,小割球を除去する実験があります。すると,残った
部分胚の大割球からは骨片形成細胞も原腸も生じました。つまり,大割球は(おそらく少量
含んでいる Dsh のおかげで)骨片形成細胞になる発生能を備えているのですが,正常発生に
おいては小割球からの抑制的な相互作用のせいで骨片形成細胞を生み出さないようになって
いるのです。小割球を切り外してしまうと,その抑制が解除されるために,大割球から骨片
形成細胞があらわれるのだと考えられます。
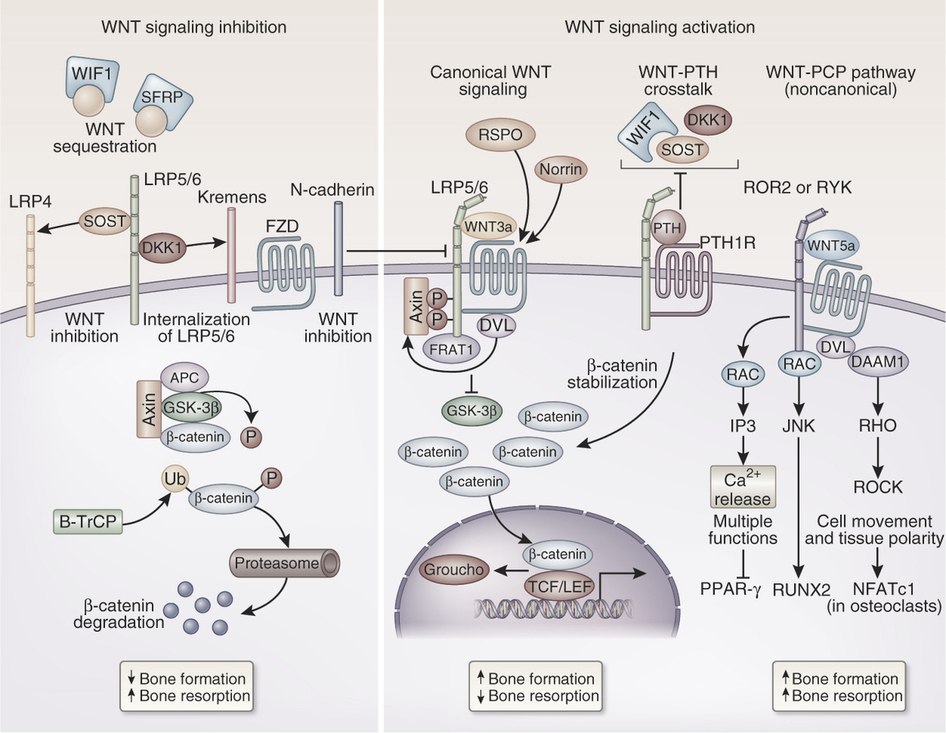
(磯江さん) 授業の中で Wnt について少し触れていましたが,詳しくはどんな働きをして
いるんですか。
〜似たような趣旨の質問: 粟田君
(答) それは,話せば恐ろしく長くなるので話しません(笑)。下図は Nature という雑誌に
あった Wnt シグナル経路の概略図です。図の中では Dishevelled が「Dsh」ではなく「DVL」
と表記されています。みんなは,グルカゴンの受容体が細胞内のタンパク質の働きをどのように
調節するかを図解したスライドを憶えていますよね。それを思い出しながら下図を見てください。
た〜っくさんのタンパク質が Wnt の影響を受けて,細胞の中で活性化したり抑制されたりして
います。詳しくは「Wnt」というキーワードでネットの検索をして見てください。数え切れない
ほどの記事が出てきますよ。
(永野君) 動物極,植物極とはどうしてこう呼ばれるのか。
〜同様の趣旨の質問: 藤木君
(答) 減数分裂では,4 つの娘細胞ができますよね。卵母細胞が減数分裂をして卵ができる
ときには 2 回の分裂ともにものすごい不等分裂が起こり,4 つのうち 1 個だけが卵となり,
残りはちい〜さな細胞(極体といいます)になります。1 回めの分裂で生じた極体は,
2 回めの分裂をしない場合が多いです。この極体の出てくる場所が動物極,その反対側が
植物極です。両生類のように大きな卵では,核は動物極側に,卵黄は植物極側に偏っています。
そのため細胞分裂のための分裂溝(卵割溝)が核の場所に近い側(動物極側)で早めに
生じて,植物極側では遅れて生じるのです。それで活発に活動しているように見えるから
“動物”極側,活動が鈍くて静かな方が“植物”極側と呼ばれるようになったのです。
でも,実際にはいろいろと重要な物質が局在していて,胚の体づくりに中心的な役割を
演じているのはどちらかというと植物極側なんですけどね。
(永野君) Dishevelled はなぜ植物極側に偏っているのか。
〜似たような趣旨の質問: 堀川さん
(答) いい質問ですね。通常,タンパク質や mRNA が細胞内で局在する場合には,微小管や
細胞骨格アクチンなどの細胞骨格が重要な役割を担っています。なので,特定の細胞骨格の
重合を阻害する薬剤…微小管の場合にはコルヒチンやタキソールやノコダゾール,細胞骨格
アクチンの場合にはサイトカラシン B やサイトカラシン D などを使います。ウニでは,
卵母細胞が成長する過程で Dsh タンパク質は植物極側に偏り始めます。この過程には微小管
が関わっていると考えられています。また植物極側の表層にある Dsh は,サイトカラシン B
やサイトカラシン D で処理すると,表層から離れて卵母細胞の内部の方へ滲んでくるので,
表層から動かないように固定する役割を細胞表層のアクチンが担っていると考えられています。
線虫 C. elegans の胚発生
(植本さん) スライド 20 を見る限り,「どちらも ABp」じゃなくて「ABa と ABp」と書いてる
ように見えるのですが,呼び方は,ABa は働きが ABp になっても ABp と何で言わないん
ですか?
(答) 下図ですよね? 図で「ABa」と書いてあるのは「本当なら ABa になるはずだった細胞」
という意味です。その方が,どっちが「元々 ABp になるはずだった細胞」で,どっちが「本来
なら ABp にはならなかったはずの細胞」か,区別がつきますからね。

(加藤君) ABa も ABp も ABp の細胞分裂をすると,どうなるのですか。ABp のところが
2 倍になるのですか。
(答) そうですね。本来 ABp から生じるはずだった組織の細胞が 2 倍になって,ABa から
生じるはずだった組織の細胞がない…という状態になるでしょうね。
(土井さん) EMS 割球は P2 と接しているので,ABp と同じ運命をたどるのですか?
(粟田君) P2 の細胞膜上にある apx-1 は EMS 側にシグナルを送ることはないのですか。
(答) それはとても重要な質問です。スライドをもう一度見直してくれたらわかりますが,
EMS 割球の系統の細胞系譜は ABp 系統とは全然違いますよね。それぞれの割球から生み
出される組織の種類も全然違います。 EMS 割球は glp-1 遺伝子を発現しておらず,
P1 や P2 からの apx-1 遺伝子産物からの誘導を受けることもできません。
(谷岡さん) apx-1 シグナル分子を受け取る glp-1 受容体を ABp だけでなく ABa も持っている
のはどういう意味があるのですか。位置が変わってしまっても大丈夫にするためですか。
(答) そうですね。持っていることには「意味」はないと思います。でも,結果的に
谷岡さんの言うように「安全装置」として役立っていると言えるかも知れません。
モザイク卵と調節卵
(永野君) モザイク卵となるか調節卵となるかは何が異ならせているのか。
〜似たような趣旨の質問: 小梶君
(答) 動物の種によって発生の仕組みはいろいろに異なっています。また,一つの卵の中でも,
決定の起こる発生段階は組織によっていろいろです。どのような物質を卵のどこに局在させ,
その物質が,細胞の活動にどの時期にどのような影響を及ぼすか…などはケース・バイ・ケース
なのです。
そんな中で,哺乳類の卵は極めて高い調節性を示します。細胞が数千個になってもまだ,
一つ一つの細胞の発生運命はそれほど限定されておらず,他の細胞に分化する能力を残して
います。初期胚の細胞には,今日の Dsh のように,局在するタンパク質は発見されておらず
一番最初のステップから細胞間相互作用によって運命が限定されていくというシステムを
採用しています。つまり,未受精卵の中に局在する物質が,特定の割球にだけ受け継がれる
という仕組みではなく,細胞分裂しているうちに,特定の細胞がたまたま特定の位置に来た
とき,特定の運命へと進むような誘導を受ける…という感じです。
哺乳類にも Dsh はあります。ただ,哺乳類の胚では局在してはいないのです。Dsh は,
ホヤや両生類の卵内でも植物極側に局在しています。ただし,Dsh を一番多量に受け取る
植物極付近の細胞は,ウニの場合には骨片形成細胞という中胚葉性の細胞になり,ホヤの
場合は内胚葉性の細胞になります。両生類の胚では,Dsh は受精の直後に,精子侵入点の
反対側に向かって移動し,濃縮されます。そして Dsh が濃縮された側が,胚の背側に
なります。Dsh はいろいろな動物の初期胚で,胚の細胞の間の“違い”を生み出すために
役立ってはいるものの,その役割は動物の種によっていろいろですね。これって,すぐ上の
平野さんの質問に対する一つの答えになってませんか?
(佐藤さん) 人間で一卵性の双子ができる仕組みは,ウニの受精卵の 4 細胞胚の細胞をバラバラに
するとそれぞれから完全な形の幼生が生じる仕組みと同じですか。また,そうなら,人間の受精卵
は調節卵であると言っていいのか。
(答) そのとおりです。一卵性の双子がどのような仕組みで生まれるのか,実際のところはよく
わかっていないのですが,本来 1 人の体を作るはずの 1 個の受精卵を出発点として,どこかの段階
で胚が 2 つに別れて,1 人の体を作ったことには間違いありません。これは調節卵だからこそ可能
なことです。
(谷井さん) ウニは 4 細胞胚の細胞をバラすとそれぞれが幼生になり,カエルは 2 細胞胚をバラ
すと右半身と左半身に分化するという話がありましたが,細胞をバラしても正常に発生するか
どうかの段階は生物の種によって異なっているということですか。
(小梶君) Driesch のウニ胚の実験の前年にあったカエルとホヤの実験から推察するに,カエルと
ホヤはモザイク卵と考えて大丈夫ですか?
(答) 谷井さんの言う「細胞をバラしても正常に発生するかどうかの段階は生物の種によって
異なっている」という結論は正しいです。そのとおりです。それはそれとして…
カエルの実験はちょっと違うのです。Wilhelm Roux が行った実験は,2 細胞期の胚の片方の
細胞を針で焼き殺す実験でした。割球をバラしてはいないので,死んだ細胞がくっついたまま
だったのです。残った 1 個の細胞から半身しかできてこなかったのは,その死んだ細胞が
あったからです。2 個の細胞をきれいにバラバラにしてしまえば,2 個ともが完全なオタマ
ジャクシになれるのですよ。カエルは一般には調節卵と言われます。ホヤは典型的なモザイク卵
です。上の方の Yang 君の質問・回答と深く関連しているので,あわせて読んでみてください。
(有吉さん) 発生能と予定運命が等しくなるとありますが,人工的に手を加えた場合は,発生能
は何と等しくなるのですか?
(答) 初期胚では,発生能は予定運命より幅広いです。例えば,ある動物の初期胚の細胞 A の
予定運命は神経だけれど,その細胞の発生能としては「表皮にも神経にもなれる」としましょう。
手を加えないで,正常に発生させた場合,その後何回かの細胞分裂を繰り返すと,A の子孫細胞
たちは,神経になることを決め,もはや神経以外の細胞(表皮細胞)には分化できない状態に
なります。これが運命の決定です。A 細胞を,初期胚の段階で他の細胞と切り離したり,別の
場所に移植したりすると,表皮になったりします。つまり,発生能として持っていたいくつかの
レパートリーの中で,予定運命とは異なる細胞になるということです。ただ,この A 細胞は
単離したり移植したりしても,神経と表皮以外の細胞にはなりません。万一,A 細胞に実験操作
を加えて筋肉細胞に分化したとしたら,A 細胞の発生能は「神経と表皮と筋肉」と書き換えないと
いけないことになりますね。
(藤木君) ウニの 4 分の 1 胚は 4 匹のプルテウス幼生になったのに,ホヤの半胚は半分になった
のですか? ホヤの受精卵は予定運命と発生能が等しくなるのが早いということですか?
(答) 実際には結構複雑なんですが,まあだいたいそんなものと思っていてくれて OK です。
生
物学概論 II のトップページに戻る