9. 変異タンパクの精製
はじめに
この実験で作ったプラスミド(pQE30)は、挿入 cDNA がコードするタンパクを簡単に大量に
発現させるために巧妙にデザインされたものであることが、前の章までの実験でわかった
と思います。しかし、このプラスミドの便利さはまだまだ他にもあるのです。日程の都合上、
皆さんが実際に作業することはできないかも知れませんが、ここで紹介するのは
大腸菌に発現させたリコンビナントタンパクをワンステップで簡単に精製する手法です。
このような方法で合成されるリコンビナントタンパクの量は、確かに、大腸菌が本来持って
いる全部のタンパクを足したよりも多いくらいです。しかし、それでも SDS-PAGE の結果から
もわかるように、皆さんがほしいタンパクは、(それぞれの量は少ないとはいえ)数え切れない
大腸菌タンパクの中に混ざって存在しているというのも事実です。例えば、皆さんはリコンビ
ナントタンパクの機能を詳細に解析したいとします。そのとき、もし、大腸菌の中に、その
タンパクとよく似た・・・しかし何十倍、何百倍の強い活性をもつタンパクがあったとしたら、
例え量が少なくてもそのタンパクの存在は無視できなくなり、実験結果も信用できないものに
なります。そこで、できることなら、目的のタンパクだけを精製しなければなりません。
今回使った pQE-30 のようなプラスミドは、作ったタンパクを精製するために便利な工夫が
ほどこされています。
知っておかなければならないこと
前の章で見た配列をもう 1 度見てみましょう(図 1)。前の実験で合成したリコンビナント
EGFP タンパクは、正味の EGFP の配列だけでなく、N 末端に余分のアミノ酸配列が付加
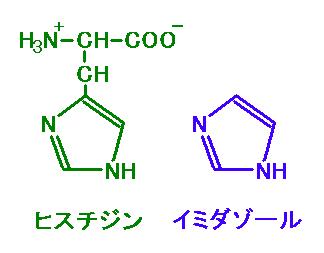
されています。ここで重要なのは 6 個ならんだヒスチジン(H)です。
図 1
この 6x ヒスチジンリピートが、リコンビナントタンパクを精製するための便利な道具なのです。
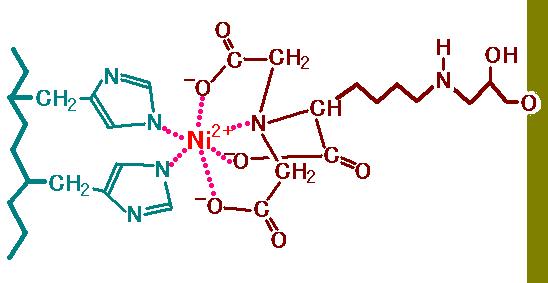
図 2 を見てください。右側にうぐいす色の壁があります。これは、固相をあらわしています。
実習で使うとすれば、この部分はアガロースゲルです。固相から飛び出している枝の先(左端)
の部分を Nitrilotriacetic acid (NTA) といいます。NTA
は、ニッケルイオン(Ni2+)の
6 つの結合箇所の 4 ヶ所を捕まえています。つまり 2 ヶ所がフリーです。その残った 2 ヶ所に
ヒスチジンが結合するのです。図では左側の青いジグザグの太線がタンパク質のペプチド
骨格をあらわしていて、そこから 2 個のヒスチジン側鎖が出ている様子を示してあります。
図 2 タンパクのヒスチジン側鎖が NTA と結合する様子
この性質を利用したアフィニティークロマトグラフィーによって、リコンビナントタンパクを
簡単に精製できるのです。N 末端に付加されたヒスチジンリピートは、たくさんの荷物の中
から自分の荷物を見分けて選び出す荷札(タグ)にたとえて、ヒスチジンタグ (His-タグ)
と呼ばれます。 His-タグは、タンパク全体の立体構造を壊すほど大きくないので、多くの
場合、タグをつけたまま活性の測定などの実験に使うことができます。もしも、タグが
そのタンパクの機能をじゃまするような場合には、タグを C 末端につけることもできます。
この実験に使った pQE シリーズの他にも、タンパク発現・精製用のプラスミドはたくさん
あります。例えば、Pharmacia から売られている pGEX ベクターは、約
26 kDa の
グルタチオン-S-トランスフェラーゼ(GST)の翻訳領域の下流に自分の
cDNA を
組み込んで、融合タンパクとして発現させるシステムです。この場合、融合タンパクは、
GST がグルタチオンと結合する性質を利用したアフィニティークロマトグラフィーによって
精製することができます。精製したタンパクは融合タンパクのまま活性測定等に使う
こともできますし、GST 部分と切り離すこともできます。GST 翻訳領域の下流、cDNA
との連結部のすぐ上流にトロンビンというプロテアーゼの認識配列 [
LVGRPS ] が
あって、そこで切り離すことができます。また、New England Biolabs で売られている
pMAL ベクターは約 42 kDa のマルトース結合タンパク(MBP)との融合タンパクを
作ることができます。この場合は MBP がマルトース(麦芽糖)と結合することを利用した
アフィニティークロマトグラフィーで精製できます。また精製後に MBP の下流にある
Factor Xa (これも血液凝固系で働くプロテアーゼです)認識部位
[ IEGR ] を切断
することによって、MBP 部分と切り離すことができます。
実験
1. 前の章と同様に IPTG でタンパク発現を誘導した大腸菌をエッペンに
移し、遠心して集菌する。菌の沈殿を残し上清を捨てる。
グループで 1 サンプル。2. エッペンに バッファー B (a) を 1 mL 注ぎ、ボルテックスで混ぜる。
3. 10 mL 用チューブに移し、超音波破砕機で合計 3 分ほど sonication。
1 回に 1 分程度、氷冷しながら行う。30 秒ほど休んで、再び超音波・・・を
繰り返す。大腸菌が破壊されれば、液が透明になる。4. 溶菌液を新しいエッペンに移し、20‐30 分間、冷却遠心する。
5. 遠心の間に、Ni2+‐NTA‐アガロースビーズをバッファー B で平衡化する。
(1) ビーズを 50 μL エッペンにとり、まず、ビーズを懸濁してある液を捨てる。
(2) 500 μL のバッファー B を加える。
(3) マイクロピペッタ−で混ぜた後、低速で遠心し、上清を捨てる。
(4) 再びバッファー B を 500 μL 加え、混ぜる。
(5) 低速で遠心し、上清を捨てる。(上清を捨てるのは、溶菌液の遠心が
終わる直前でよい)6. 溶菌液の遠心が終わったら、上清のうち 50 μL を別のエッペンにとり
等量の 2x SDS-サンプルバッファーと混ぜる。残りの上清を平衡化済み
のビーズの入ったエッペンに移す。
液はたくさん残してもいいので、沈殿を絶対にとらないように。7. 30 分ほど室温に置き、ときどきピペッタ−で混ぜる。
Ni2+‐NTA‐ビーズに His-タグ付きリコンビナントタンパクを吸着させる。8. 低速の遠心でビーズを沈殿させた後、上清を捨てる。
9. ビーズに バッファー C (b) を 1 mL 加え、混ぜる。
バッファー C はビーズの洗浄用。このバッファー中では His‐タグタンパクは
ビーズから離れない。ビーズの隙間などに(Ni2+-NTA に吸着したわけでも
ないのに)残っている不要なタンパクを除去する。10. 手回し遠心でビーズを沈殿させ、上清の 50 μL を別のエッペンに
移し、等量の 2x SDS‐サンプルバッファーを加える。
残りの上清を捨てる。11. ステップ 9 と 10 をもう 1 度繰り返す。
ただし、SDS-PAGE 用のサンプリングはしなくてよい。12. ビーズに バッファー E' (c) を 100 μL 加え、混ぜる。
バッファー E' には イミダゾール(d) が入っている。これによって、His-タグ
タンパクと Ni2+-NTA との結合が阻害され、タンパクはビーズから離れる。
13. 室温で 5 分ほど置き、ときどき混ぜる。
14. 低速の遠心でビーズを沈殿させ、上清を新しいエッペンにとる。このうち
50 μL を別のエッペンにとり、等量の 2x SDS‐サンプルバッファーを
加える。15. ステップ 12-14 をもう 1 度繰り返す。
ただし、SDS-PAGE 用のサンプリングはしなくてよい。16. SDS‐サンプルバッファーにとかした各サンプルを 80-90℃ で 5 分
煮て、15-20 μL 程度を電気泳動する。
実験に使う試薬
8 M 尿素(b) バッファー C
100 mM リン酸二水素ナトリウム(NaH2PO4)
10 mM Tris-HCl (pH 8.0)
8 M 尿素(c) バッファー E’
100 mM リン酸二水素ナトリウム(NaH2PO4)
10 mM Tris-HCl (pH 6.3)
8 M 尿素(d) イミダゾール
100 mM リン酸二水素ナトリウム(NaH2PO4)
10 mM Tris-HCl (pH 8.0)
250 mM イミダゾール (下を参照)
図 3 下図の左がヒスチジン、右がイミダゾール。図 2 と見比べよう。
イミダゾールが大量に持ちこまれると、このイミダゾールが Ni2+-NTA と結合し、
その結果、His‐タグタンパクがビーズから離れる。