2‐2. 全 RNA からの cDNA 合成
(逆転写)
はじめに
全 RNA から、cDNA の合成(逆転写)を行います。2‐1 で抽出・精製
した RNA サンプルの 80% 以上は rRNA であり、残りの大部分は tRNA です。
mRNA は全 RNA 中の数パーセントに過ぎません。しかし、真核生物の mRNA
のほとんどには、3’末端に(遺伝子 DNA にはもともとなかったポリ (A) 配列
(アデニンヌクレオチドが数十から数百塩基ならんだ配列)が付加しています。
この特徴的な配列を利用して、mRNA に相補的な DNA (これを
complementary DNA = cDNA といいます)を合成することができます。
まずはチミンヌクレオチドが並んだ短い DNA 鎖を用意します。今回は、
次の作業に便利なように、
[5'-GACTCGAGTCGACATCGATTTTTTTTTTTTTTTTT-3']
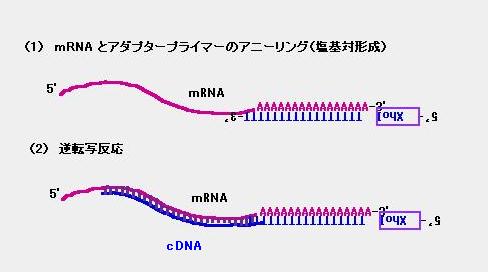
という配列の DNA 鎖を用意します。これを mRNA と混ぜると、mRNA 持つ
ポリ (A) 鎖との間でアニーリング(塩基対の形成)が起こります [図 1 の (1)]。
その後、逆転写酵素を加えると、 mRNA を”鋳型”として、mRNA の配列と
相補的な DNA (cDNA) が合成されるわけです [図 1 の (2)]。
図 1
知っておかなければならないこと
一般的に、DNA の鎖が合成されるときには、合成開始場所に短い DNA あるいは
RNA の鎖が最初に塩基対を作っている必要があります。DNA の鎖は、あらかじめ
塩基対を作っている鎖の 3' 末端にヌクレオチドを付加するように起こります。
このように、DNA 合成の”足場”になるような短い鎖をプライマーと呼びます。
mRNA に特徴的なポリ (A) 配列に対してアニーリングするようにデザインされた、
チミンヌクレオチドの連続からなる DNA 鎖を一般にオリゴ(dT)プライマー
と呼びます。今回使用するプライマーは、5' 末端に余分の(mRNA とは
アニーリングしない) [ GACTCGAGTCGACATCGAT ] という配列をつけてあります。
この配列の意味は 2-4 で明らかになります。今回用いる、このようなプライマーを
ここでは dT-アダプタープライマーと呼ぶことにします。
通常の細胞の活動において DNA が合成されるのは、細胞分裂周期の”S 期”
すなわち DNA 合成期です。このとき働く酵素は DNA を鋳型にして、それに
相補的な DNA 鎖を合成する DNA 依存性 DNA ポリメラーゼです。普通に
DNA ポリメラーゼと言われるのはこのタイプの酵素です。ほとんどの DNA
ポリメラーゼは、RNA を鋳型にして DNA を合成する活性をもっていません。つまり、
酵素が DNA のみを鋳型として認識するわけです。では、今回の実験で、mRNA を
鋳型としてこれに相補的な DNA を合成するにはどうしたらよいのでしょうか?
ここで役に立つのが逆転写酵素です。真核生物に感染するウィルスの中には
AIDS ウィルスなどのようにウィルスの遺伝子が RNA でできているものがあります。
要するにウィルス粒子の殻の中に RNA が遺伝子として詰めこまれているわけです。
ウィルスは細胞に感染し、遺伝子 RNA を細胞に送りこみます。しかし、ウィルスの
遺伝子が発現するためには、やはり RNA のままではダメで、ウィルスは何より
先にその RNA を鋳型として DNA を合成しなければなりません。このとき働くのが
ウィルスの殻の中に仕込まれていた逆転写酵素なのです。逆転写酵素は、
RNA 依存性 DNA ポリメラーゼです。この後、合成されたウィルスの
DNA
から様々なタンパクをコードする遺伝子が発現しはじめるわけです。
今回使う逆転写酵素はマウスの白血病ウィルスから単離されて商品化されている
Moloney Murine Leukemia Virus Reverse Transcriptase
(MMLV RTase)です。
さて、この実験で合成する cDNA は mRNA に相補的な配列をしています。
では、この配列は遺伝子の配列とはどう違うのでしょうか?真核生物においては
遺伝子が転写されてできる最初の RNA (昔は heterogeneous nuclear RNA と
呼ばれていた)には、ほとんどの場合、成熟 mRNA にはない配列が含まれて
います。これをイントロンといいます。この配列は、スプライシングと呼ばれる
反応によって切り捨てられます。また、3' 末端にはポリ (A) 配列が、5' 末端には
キャップ構造(7-メチルグアノシンがその5' 末端を RNA の 5'
末端と向かい
合わせにして・・・つまり逆向きに・・・結合したもの)が付加されます。真核生物の
リボソームは、一般的にこのキャップ構造を認識して会合します。したがって
mRNA を鋳型として合成した cDNA には、イントロンの配列は含まれず、
ポリ (A) 配列を持つことになります。
なお、真核生物でも、DNA と結合して染色体を構成するヒストンなど
ハウスキーピングな(細胞の生存に必要な)タンパクをコードする mRNA
の中に
はポリ (A) 配列を含まないものがあります。オリゴ (dT) プライマーを利用して
cDNA を合成するときにはそのような例外は無視しているわけで、この実験の
説明文のあっちこっちで「mRNA に相補的なDNA を合成…」と書いて来ましたが、
正確に言えば、ここで行う実験は ポリ (A)+ RNA
を鋳型として、それに相補的な
DNA を合成する作業です。
実験
1. 前の実験で抽出・精製した全 RNA を 5 μl 新しいエッペンにとる。
注 1: 本来は RNA 量を測定してから使用するべきであるが、実習では
省略。定量してから使用する場合は、全 RNA が 1 μg あれば、
比較的レアな(細胞あたり 1 本から 10 本程度しかないような)
RNA からでも増幅が可能であると言われている。
注 2: cDNA ライブラリー(分子生物学、遺伝子工学などの講義で
習うかな?)を作る場合などには、ポリ (A)+ RNA のみを精製して、
純度の高いポリ (A)+ RNA を数マイクログラム使用しなければ
なりません。2. dT-アダプタープライマー (a) を 3 μl 加えて、混ぜる。
注: このように、少量の液を測り、混ぜるときは、非常に小さな 1 滴の
溶液が大きな誤差の原因になる。
マイクロピペットの使い方についての注意点は後述。3. 90℃ で 3 分加熱後、素早く氷冷する。
RNA は 1 本の鎖でできており、回文(palindrome)配列
(例えば AAAGGGGCCCCTTT というような)などがあると、同一分子内
で塩基対(2 次構造という)を作ることがある。特に、mRNA の
3' 側非翻訳領域には、機能的に重要な 2 次構造がある場合が多く
(細胞工学の講義を聴こう)、cDNA 合成の際に、その箇所で DNA
合成がストップしてしまうことがある。そこで、加熱によって塩基対
(水素結合)をはがし、つづく急冷によって分子の運動を小さくし、
再び 2 次構造が形成されるのを防ぐ。4. 軽く遠心して、液をエッペンの底に落とす。
加熱時に蒸発しフタに結露した液を、遠心で底に落とす。上の
ステップ 2 の注意事項で述べたことと同様で、わずかの液の蒸発も
無視できない誤差の原因になる。5. 逆転写反応液を調製する。
以下の試薬を順に加える。
5x 逆転写バッファーを 4 μl
[250 mM Tris-HCl (pH 8.3), 375 mM KCl, 15 mM MgCl2]
100 mM DTT (c) を 2 μl
dNTP mixture を 4 μl
[4 種類のデオキシリボヌクレオシド三リン酸(dNTP)を
各 5 mM になるように溶かしたもの]
20 units/μl RNase inhibitor (d) を 1 μl
200 units/μl MMLV RTase (e) を 1 μl6. 反応液を 37℃ のウォーターバスに 1 時間おく。
7. TEバッファー (b) を 80 μl 加える。
これを、次の実験(PCR によるヘモグロビン cDNA の増幅)に用いる。
逆転写が終わった後は、RNA は分解しても構わない。むしろわざわざ
分解してから次のステップに進む人もいるくらいである。ここから先は
DNA を扱うことになる。
実験に使う試薬
[5'-GACTCGAGTCGACATCGATTTTTTTTTTTTTTTTT-3'](b) TE バッファー
という配列の 1 本鎖 DNA を 10 pmol/ml になるように TE バッファーに
溶かしたものを使う。(TTTTTTTTTTTTTTTT)の部分が、ポリ (A) 配列と
塩基対を作る。
注: TE バッファーの組成は以下のとおり。
10 mM Tris-HCl (pH 7.5 あるいは 8.0 が普通)、1 mM EDTAを溶かした(c) DTT (dithiothreitol)
水溶液。調製後にオートクレーブする場合はフタを完全に閉じて
おかないと、塩化水素が飛んで pH が上がるので注意。
DNA を分解するような活性のある酵素の多くは、反応に Mg2+ イオンを
要求するため、2 価イオンのキレート剤である EDTA を含むバッファー
中では働きにくい。TE バッファーにとかした DNA は、冷蔵庫で何年でも
安定に保存できる。
還元剤。逆転写酵素や RNase inhibitor は DTT 存在下でないと(d) RNase inhibitor
活性が低い。
1 本鎖 RNA を分解する RNA 分解酵素の活性(RNase A 活性という)を(e) MMLV RTase
阻害するタンパク。今回使用するのは、ウシのすい臓から精製された
RNase inhibitor (東洋紡)。このタンパクは RNase と 1 対 1 の割合で
結合して、その活性を阻害する。この他、非タンパク質性の RNase
阻害剤もあるが、それらの多くは逆転写反応にも悪影響を及ぼすことが
多いので最近は使われない。
詳しい説明はこのページの「知っておかなければならないこと」の中で
説明済み。今回使うのは Gibco BRL 社製。この他にトリの肉腫ウィルス
からとられた逆転写酵素なども広く使われている。
一般的に、逆転写酵素は RNA を鋳型としてそれに相補的な DNA を
合成しつつ、鋳型となった RNA (合成後は DNA と RNA の 2 重らせんを
形成している)を分解する活性(RNase H 活性という)をもつ。RNase H
活性とは DNA と 2 重らせんを形成している RNA のみを分解する
活性であり、 RNase A 活性とは異なる。逆転写反応においては、
RNase H 活性が強すぎると合成途中の DNA と鋳型 RNA との結合が
弱くなって、合成が途中で終わってしまうことがある。そこで MMLV
RTase を遺伝子工学的に改変して RNase H 活性をなくしたものも
市販されている。