
レチノイン酸の標的遺伝子
レチノイン酸受容体は、DNA に結合し遺伝子の転写を活性化する転写調節因子です。
芽体の多能性細胞(囲鰓腔上皮)を、レチノイン酸で直接処理しても何も起こりません。
しかし、レチノイン酸で処理した間充織細胞を、(レチノイン酸を洗い流した後に)芽体に
移植すると二次軸を誘導することができます。このことから、レチノイン酸が直接働き
かける相手は間充織細胞であり、間充織細胞がレチノイン酸に応答して分泌する分子
が囲鰓腔上皮に直接働きかけて分化転換を誘導することが示唆されます。
私たちは、間充織細胞をレチノイン酸処理したときに活性化する遺伝子をディファレン
シャルディスプレイ法という方法で単離しました(図
1)。(この研究では東大医科研の
伊藤隆司先生、鈴木高史先生にお世話になりました。)
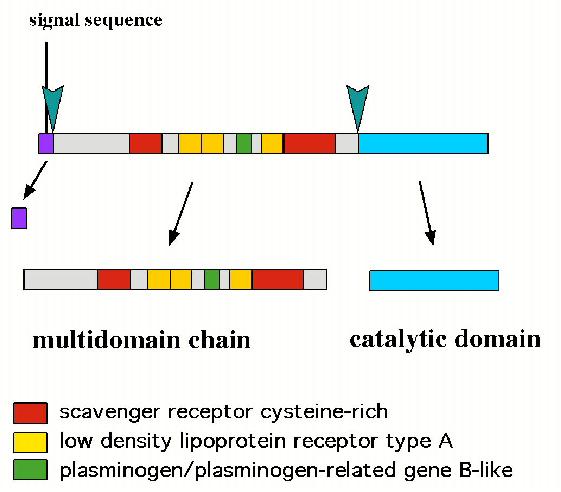
単離された遺伝子のうちの一つは複雑な構造をしたセリンプロテアーゼでした(図
2)。
芽体では、プロテアーゼの活性が上昇していること、未精製のプロテアーゼを芽体に
注入すると二次軸ができることなどが、川村らによって明らかにされていることから、
このプロテアーゼが囲鰓腔上皮の分化転換を誘導する因子ではないかと考えています。
現在、このプロテアーゼの機能解析を行っています。川村らは、特にセリンプロテアーゼの
触媒ドメインに着目して、培養細胞を用いた機能解析を行っています。
また、このプロテアーゼ以外にも、ゴナドトロピン放出ホルモン受容体と思われる
7 回膜貫通型の G タンパク共役型受容体など、複数の
cDNA が見つかっています。
これらの cDNA がコードするタンパクの機能解析も重要な課題です。
図 1 (左): ディファレンシャルディスプレイの結果。レチノイン酸処理をした間充織
細胞(+)で発現していて、処理していない細胞(-)で発現していない遺伝子を探した。
図 2 (右): ディファレンシャルディスプレイで同定された
cDNA のうち一つは
プロテアーゼをコードしていた。N 末側には、他のタンパクと結合すると思われる
モチーフが多数見られ、C 末側にセリンプロテアーゼの触媒ドメインがあった。
参考文献
Ohashi, M., Kawamura, K., Fujii, N., Yubisui,
T. & Fujiwara, S. (1999)
A retinoic acid-inducible modular
protease in budding ascidians.
Develop. Biol. 214:
38-45.
川村和夫 ・ 藤原滋樹 ・ 指吸俊次 (2000) ホヤの無性生殖とプロテアーゼ
〜細胞増殖・脱分化における TRAMP
の役割 生化学 72: 298-302.
Fujiwara, S., Kamimura, M., Ohashi, M. &
Kawamura, K. (2001) Molecular bases
of bud development in ascidians.
In
The Biology of Ascidians. (ed. Sawada, H.,
Lambert,C. & Yokosawa,H.) Elsevier
Science Publisher, pp.300-304.