レチノイン酸受容体
レチノイン酸は水に溶けにくく油に溶けやすい(脂溶性の)物質です。その受容体は
細胞の表面ではなく核の中にあります。受容体は、レチノイン酸受容体(Retinoic
acid
receptor, RAR) とレチノイド X 受容体(retinoid
X receptor, RXR)という 2 種類の
タンパクが結合したもの(ヘテロ二量体 = heterodimer)です。この受容体については
脊椎動物ではよく研究されていて、配列特異的に
DNA に結合し、レチノイン酸と結合
すると転写活性化因子として働くということがわかっています。
RAR は、脊椎動物から
しか見つかっていませんでしたが、私たちは、無脊椎動物としては初めて、ミサキマメ
イタボヤから、RAR の cDNA を単離しました。さらに、ホヤの
RAR/RXR ヘテロ二量体
が、配列特異的に DNA に結合すること(下図)、またレチノイン酸依存的に遺伝子の
転写を活性化することを証明しました。
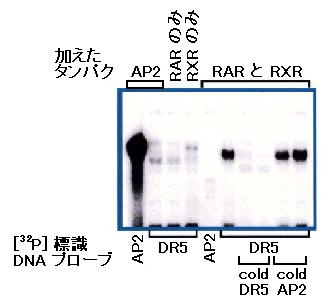
図: ゲルシフト解析の結果。ゲルシフト解析では、放射性同位元素などで標識した
決まった配列の DNA を、あらかじめタンパクと混ぜた後に電気泳動します。もしも
タンパクがその DNA と結合すれば、電気泳動での移動度が小さくなるということを
利用して、タンパクと DNA との結合を調べる実験法です。黒いバンドが出ていれば、
DNA とタンパクが結合したことがわかるという仕組みです。この実験では、RAR
のみ、
RXR のみ、RAR と RXR を混ぜたもの、そして対照実験として無関係な転写調節
因子 AP2 を用いました。一方、標識 DNA の方は
AP2 と結合することがわかって
いる配列の DNA (AP2)、脊椎動物の RAR/RXR ヘテロ二量体が結合することが
わかっている配列の DNA (DR5)を用いました。図から明らかなように、ホヤの
RAR
タンパクと RXR タンパクは、(同時に存在すれば)
DR5 標識 DNA と結合することが
わかります(右から 5 番めのレーン)。 RAR も
RXR も、単独では DR5 配列に結合
しません(左から 3-4 番めのレーン)。 RAR と
RXR は、AP2 配列には結合しません
(左から 5 番め)。一方、AP2 タンパクは AP2 配列には結合しますが(左端)、DR5
には結合しません(左から 2 番め)。右側の 4 レーンでは、標識
DNA とタンパク(RAR
と RXR)の反応液に大量の非標識 DNA (cold DNA)を混ぜました。多量の非標識
DR5 配列は標識 DNA とタンパクを奪い合うため、結果としてバンドの放射活性は
弱くなります。一方、非標識 AP2 配列をどれだけ加えても、DR5
と RAR/RXR との
結合を競合阻害しないので、バンドは弱くなりません。
参考文献
Hisata, K., Fujiwara, S., Tsuchida, Y., Ohashi,
M. & Kawamura, K. (1998)
Expression and function of a retinoic
acid receptor in budding ascidians.
Develop. Genes Evol. 208:
537-546.
Kamimura, M., Fujiwara, S., Kawamura, K. &
Yubisui, T. (2000) Functional
retinoid receptors in budding ascidians.
Dev.
Growth Differ. 42: 1-8.
Fujiwara, S., Kamimura, M., Ohashi, M. &
Kawamura, K. (2001) Molecular bases
of bud development in ascidians.
In
The Biology of Ascidians. (ed. Sawada, H.,
Lambert,C. & Yokosawa,H.) Elsevier
Science Publisher, pp.300-304.