私たちの研究の背景
出芽の説明のページ で見たとおり、ミサキマメイタボヤの芽体は、親から離れてから
体づくりを始めます。親から離れた芽体では、親に一番近い側の囲鰓腔上皮の分化
転換によって消化管ができます。
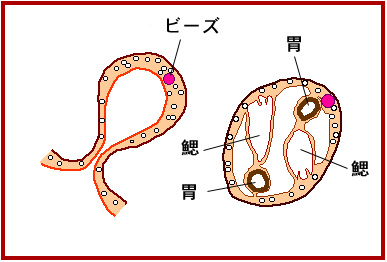
ところが、芽体の、親から遠い側にレチノイン酸(ビタミンAからできる)を染み込ま
せたビーズを埋め込むと、親に近い側にできる本来の消化管に加えて、ビーズのそば
にもう一つの消化管ができます。やがて、鰓も二つ備えた、完全な”双子”の芽体が
できます(下図)。
芽体の、親に近い側では、レチノイン酸を合成する酵素と思われるアルデヒド脱水素
酵素(aldehyde dehydrogenase; ALDH)の活性が見られます。また、親個体にはレチ
ナール(レチノイン酸はレチナールが酸化されてできる)が豊富に存在しますが、芽体
ではそれが著しく減少します。これらのことから、レチノイン酸は、単に「外から加えたら
悪い影響を与える」というものではなくて、実際の芽体の中で囲鰓腔上皮を分化転換
させて消化管をつくるために必要なものなのではないかと考えています。
参考文献
藤原滋樹・川村和夫 (1996) 芽体形成のメカニズム 遺伝第50巻12号 pp. 29-34.
Fujiwara,S. & Kawamura,K. (1992) Ascidian budding as a
transdifferentiation-like system: multipotent
epithelium is not
undifferentiated. Dev. Growth Differ.
34: 463-472.
Hara,K., Fujiwara,S. & Kawamura,K. (1992) Retinoic acid can induce
a
secondary axis in developing buds of a colonial
ascidian, Polyandrocarpa
misakiensis. Dev. Growth Differ. 34: 437-445.
Kawamura,K., Hara,K. & Fujiwara,S. (1993) Developmental role of
endogenous retinoids in the determination
of morphallactic field in
budding tunicates. Development
117: 835-845.
Kawamura,K. & Fujiwara,S. (1994) Transdifferentiation of pigmented
multipotent epithelium during morphallactic
development of budding
tunicates. Int. J. Dev. Biol.
38: 369-377.
さて、本当に、芽体でレチノイン酸が合成されて、働いているのだとすれば、
レチノイン酸合成酵素や、レチノイン酸受容体が、芽体で働いているはずです。
さらに、レチノイン酸受容体の働きによって、囲鰓腔上皮の分化転換を進めるために
必要な様々な遺伝子が活性化しているはずです。それらの遺伝子を手に入れて、
その機能を調べれば、芽体の体づくりの仕組みがわかるはずです。ひいては、
「なぜ私たちは出芽できないのにホヤは出芽できるのか」という問いに対する答えにも
近づけるのではないだろうかと考えているわけです。
以下は、その考えに沿って、私たちが進めている研究の紹介です。